
I am an NIH/NIGMS K99 Pathway to Independence fellow in Dr. Wolf-Dietrich Heyer’s lab at the University of California, Davis (previously supported through: NCI T32 CA108459-14 and the A.P. Giannini Foundation). I am broadly interested in the mechanisms that prevent and, conversely, that promote recombination-associated genome rearrangements and loss of heterozygosity (LOH) and their relationship to human health, particularly cancer and neurological conditions, such as Alzheimer’s.
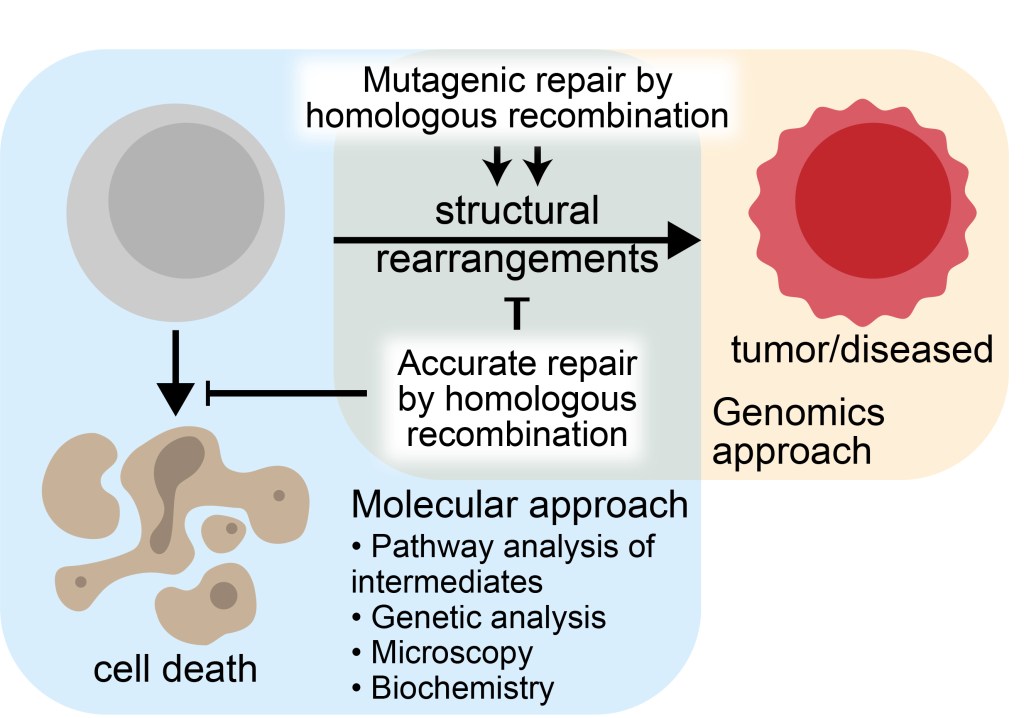
Structural rearrangements, rather than single nucleotide changes, are the predominant class of driver mutation across many cancer types. Homologous recombination is a ubiquitous and normally high-fidelity pathway to repair and tolerate DNA damage and to restart stalled or collapsed replication forks. Using the allelic site of sister chromatid as a repair template, it replaces the lost or damaged bases in a copy-paste type reaction that involves a DNA polymerase. Reversibility of multiple key steps in the pathway allows for quality control. Errors or defects in these regulatory mechanisms can lead to inaccurate repair. In particular, recombination between similar but non-identical sequences, such as repetitive elements, can lead to genome rearrangements. Moreover, errors during recombination can lead to loss of heterozygosity, which is known to drive tumor formation in patients with germline mutations in DNA repair proteins, including BRCA1/2 (breast and ovarian cancer susceptibility) and MSH2/3/6 (Lynch syndrome, also known as hereditary non-polyposis colorectal cancer or HNPCC). My current work focuses on:
- How MSH2-MSH6 prevents recombination between mismatched repeats (Heteroduplex rejection): The mismatch repair proteins MSH2 and MSH6 form a heterodimer that detects base-base mismatches and small indels. Their activities are essential during post-replicative mismatch repair (MMR) and heteroduplex rejection, a process in which a three-stranded intermediate (a so-called displacement loop, or D-loop) in the recombination pathway is disassembled if it contains as little as one mismatch. However, the precise details as to how MSH2-MSH6 mediates heteroduplex rejection, and the downstream effectors it recruits, are poorly understood. My goal is to further elucidate this important regulatory pathway in yeast and humans using a combination of genetic and biochemical approaches. I also aim to better understand how loss of heteroduplex rejection shapes the mutational landscape in MSH2/6-deficient tumors using long-read sequencing to identify de novo structural variants present in tumors relative to matched healthy tissue controls.
- Mechanism of multi-invasion-induced rearrangements: Multi-invasion recombination is a mutagenic pathway in which a single ssDNA end pairs with two or more template molecules during recombination, instead of the typical single template. Resolution of the multi-invasion pairing intermediate leads to a truncated ssDNA end and either a translocation with secondary DNA double-stranded breaks or an insertion. My current research seeks to define the differential contributions of the structure-selective endonucleases, including Mus81-Mms4, Yen1, and Slx1-Slx4, to resolving multi-invasion intermediates in order to better understand how multi-invasion-induced rearrangements form.
- Impact of 3′ heterologous flaps on recombination dynamics: Pairing of two non-allelic repeats with one another can lead not only to mismatches within the pairing intermediate, but also a 3′ heterologous flap at the end of the repeat region that prevents extension of the intermediate by a DNA polymerase. Cleavage of the flap by Rad1-Rad10 creates a 3′ end that is incorporated into the pairing intermediate and competent for extension, but may result in loss of genetic information in addition to genome rearrangements. I am using pathway and genetic analysis to determine how these structures influence the dynamics and genetic dependencies of recombination.
In addition to my work on recombination-mediated genome rearrangements and loss of heterozygosity, I am also interested in fundamental questions regarding how homologous recombination is regulated, particularly D-loop formation and extension.
I received my Ph.D. in Genetics, Genomics, and Systems Biology (GGSB) from the University of Chicago, where I worked with Dr. Douglas Bishop. My research focused on understanding the factors that regulate the meiotic recombinase Dmc1. Through my doctoral work, I came to appreciate the multitude of regulatory mechanisms that ensure that homologous recombination leads to high-fidelity repair of DNA double-stranded breaks and other sources of DNA damage.
* Banner represents EM imaging of a filament formed by RecA homolog Dmc1 on ssDNA
